

23 Août 2010

L’examen biologique des réalisations, un outil incontournable pour la gestion de l’espèce
Marc Collyn,
CNRS, Université de Rennes1, UMR 6553, Station Biologique de Paimpont.
Résumé
Il est admis que le cerf connaît une croissance régulière, jamais vue auparavant. Il
convient donc de s’interroger sur l’efficacité des actions menées et d’en tirer les
conclusions. Nous sommes capables de restaurer, voire de réimplanter des
populations, mais nous n’avons pas la maîtrise pour les réguler au seuil de densité
souhaité. Il en résulte des plans de chasse qui aboutissent trop souvent à l’explosion
démographique de ces populations, puis à leur « extermination » massive… A ce
constat, il convient d’ajouter le fait que nous ne disposons pas de statistiques fiables
sur les réalisations. L’étude suivie, depuis 10 années à Paimpont, montre que la
batterie classique de bio-indicateurs, mise en place avec l’aide des organismes
compétents, a donné des résultats contradictoires et non prédictibles. Quand les
indicateurs sont au rouge… c’est déjà trop tard car ils alertent de l’impact du cerf sur
le milieu avec un effet retard amplifié par le comportement de l’espèce. Ces bioindicateurs,
plus adaptés à la gestion des habitats forestiers, ne permettent donc pas
une gestion “durable” des populations de cerfs.
Il est également admis que « la gestion harmonieuse entre forêt et herbivores
s’articule autour de deux types d’actions complémentaires : le contrôle par la chasse
des effectifs d’ongulés sauvages et la gestion des habitats forestiers… ». Nos
travaux ont pour objectif principal de contribuer au développement de la 1ère de ces
2 actions : aider les responsables cynégétiques à contrôler les effectifs d’ongulés
sauvages par des approches cynégétique et biologique cumulées. Nous avons opté
pour le développement de 2 outils de gestion simplifiés, en impliquant les
« chasseurs », puisqu’il revient à ces derniers de contrôler les effectifs par la chasse.
Nos expériences montrent qu’ils peuvent apporter une contribution significative. Cet
effort supplémentaire sera, sans nul doute, leur meilleur investissement pour l’avenir
et le maintien de l’espèce pour le contentement de tous.
Dans le cadre de ce symposium, nous commenterons les deux outils proposés pour
la gestion de l’espèce, que nous testons plus particulièrement dans les départements
22, 35, 36, 44 et 56, avec la participation des responsables cynégétiques : i) l’apport
de l’Examen Biologique des Réalisations ; ii) le suivi de la Structure et de la
Dynamique des Biches.
Indicateurs de Changement Ecologique pour la gestion des populations de cerfs
Mathieu Garel, Michallet J, Bonenfant C, Hamann J-L, Gaillard J-M & Klein F
CNRS – Laboratoire de Biométrie et Biologie Évolutive, Lyon
ONCFS – CNERA Cervidés Sanglier, Bar le Duc
Résumé
À la fin des années 70, la mise en place d’un plan de chasse s’est généralisée à l’ensemble
des espèces gibiers. L’objectif était avant tout de permettre le maintien d’un juste équilibre
entre population et milieu. Dans ce contexte, les gestionnaires ont eu besoin de méthodes
permettant de suivre les variations d’abondance des populations de grands herbivores. C’est
ainsi qu’ont été généralisées les méthodes classiques de dénombrements, basées le plus
souvent sur des observations exhaustives. La comparaison avec des méthodes robustes et
validées dans un cadre scientifique ont cependant montré que la plupart des méthodes
utilisées par les gestionnaires sous-estimaient l’abondance réelle. Associée à un plan de
chasse protectionniste, la gestion ainsi pratiquée a largement contribué à l’explosion
démographique et géographique des populations de cerfs. Dans la phase d’abondance
actuelle, un certain nombre de problèmes commence à émerger (dégâts forestiers), qui est
d’autant plus préoccupant qu’en parallèle le nombre de chasseurs en France diminue (2-
3%/an).
Dans ce contexte, l’approche par indicateurs de changements écologiques (ICE) s’est
développée, avec pour objectif de répondre à la fois aux limites techniques–scientifiques des
approches utilisées jusqu’alors et de permettre une meilleure compréhension du système dans
son ensemble. Un ICE est un paramètre mesuré sur un animal ou un végétal dont l’évolution
est dépendante de celle du système population-environnement. Plutôt que de caractériser une
situation ponctuelle, on cherche à s’informer sur l’histoire et l’évolution du "système" pour
permettre une gestion raisonnée et adaptative, a posteriori, des populations. Derrière
l’utilisation de ces ICE, il y a en outre l’idée qu’il n’est pas possible de caractériser et
d’interpréter les changements d’état du système population-environnement à partir d’un seul
élément (comme l’abondance par exemple). L’approche par ICE n’a ainsi de sens qu’au
travers de leur complémentarité, inscrite dans un suivi temporel de longue durée. Pour statuer
sur l’état d’un système population-environnement, on reconnaît donc en général trois
catégories d’ICE qui vont nous renseigner sur (1) les variations d’abondance de la population,
(2) la performance des individus et (3) la pression de la population sur son milieu.
Nous nous proposons ici de revenir sur les principes d’utilisation des ICE dans le cadre du
suivi et de la gestion des populations de cerfs. Si peu de paramètres ont été validés en tant
qu’ICE chez cette espèce, les recherches en cours sont très prometteuses et ouvrent de
nouveaux horizons de gestion. Pour illustrer notre propos, nous prendrons comme référence
la réserve nationale de chasse et de faune sauvage de la Petite Pierre (Bas-Rhin) où l’Office
National de la Chasse et de la Faune Sauvage assure depuis plus de 30 ans un suivi
approfondi de la population présente. A la fois la qualité et la quantité des données
disponibles, nous permettront d’aborder à travers cet exemple concret les notions de
performance (poids des faons, âge de première reproduction des femelles) et d’abondance
(comptages aux phares) à la base de la “philosophie” ICE.
Retour sur une gestion par indicateurs
Contexte
La généralisation du plan de chasse des cervidés dans les années 70 avait pour objectif “le maintien
d’un juste équilibre entre le milieu, la quantité et la qualité des animaux vivants sur un territoire”. Des
méthodes de dénombrements exhaustifs ou sur secteurs échantillons (battue totale, affût et approche
combinés, poussée sur secteur échantillon et circuits voitures; Bourlière 1969, Boscardin 1999) ont
donc été vulgarisées dans l’urgence pour permettre aux gestionnaires d’estimer les effectifs de
cervidés présents sur leurs territoires. Les résultats obtenus étaient généralement complétés par une
estimation succincte des potentialités de la zone chassée et des paramètres démographiques de la
population (par exemple taux de reproduction). C’est sur cette base qu’un plan de prélèvement annuel
était défini pour maintenir une densité d’individus compatible avec l’équilibre recherché.
Au fur et à mesure de l’avancement des recherches sur les méthodes de dénombrement des
populations sauvages et l’application par certains gestionnaires des méthodes développées, il s’est
avéré qu’aucune ne reflétait correctement les réalités du terrain (Gaillard 1988). En effet, la plupart
des méthodes avait tendance à fortement sous estimer les effectifs recensés (Andersen 1953). Un tel
biais associé à la mise en place d’un plan de chasse plutôt protectionniste a largement contribué à
favoriser l’expansion géographique (figure 1) et démographique des populations de cervidés en
France (figure 2; voir aussi la communication d’Emmanuelle Pfaff lors de ce colloque)
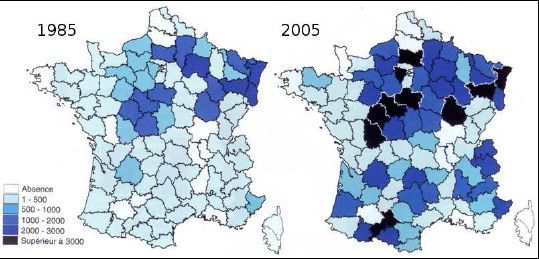
Figure 1 Exemple de l’expansion géographique des populations de cerfs. La surface occupée par l’espèce a
été multipliée par 1.6 en 20 ans.
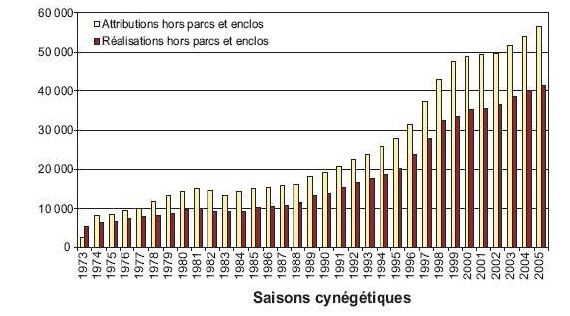
Figure 2 Exemple de l’expansion démographique des populations de cerfs. Le plan de chasse de l’espèce a
été multiplié par 4.1 en 20 ans.
Ces changements sont à l’origine d’un certain nombre de problèmes: dégâts agricoles et forestiers,
augmentation du nombre de collisions avec les usagers de la route ou encore augmentation des
risques sanitaires (maladie de Lyme par exemple). Des conflits d’intérêts entre chasseurs, agriculteurs
et forestiers sont alors apparus. Cette abondance préoccupe d’autant plus aujourd’hui qu’en parallèle
le nombre de chasseurs en France diminue: depuis 1975, la diminution enregistrée est régulière, entre
2 à 3% par an (Chardonnet et al. 2002). Dans le même temps, la population de chasseurs est
vieillissante laissant suggérer à terme une diminution encore plus rapide (l’âge moyen des chasseurs
a augmenté de cinq ans en quinze ans, passant de 45 à 50 ans, Lecocq & Meine 1998). Nous
pourrions donc être dépassés par l’ampleur des changements qui affecte notre écosystème en terme
d’espèces herbivores. Ceci soulève évidemment de nombreuses questions comme la compétition
entre espèces (cerf et chevreuil par exemple) qui ne partageaient pas jusqu’à présent leur habitat (voir
aussi la communication d’Emmanuelle Richard lors de ce colloque).
L’alternative ICE
Dans cette phase d’abondance actuelle, il est devenu indispensable de trouver des alternatives aux
méthodes de dénombrement. De telles alternatives doivent notamment permettre de résoudre les
problèmes de fiabilité, mais aussi de lourdeur d’organisation et de coût élevé associés aux méthodes
traditionnelles qui limitaient fortement la surface échantillonnée.
Devant cette nécessité, les organismes en charge de programme d’étude sur le fonctionnement des
populations de grands mammifères (CEMAGREF, CNRS, INRA et ONCFS) se sont appliqués à
définir une stratégie de gestion basée sur l’utilisation d’une batterie d’indicateurs de changement
écologique (ICE; Morellet et al. 2007). Toute variable qui décrit l’état d’une population en relation avec
son habitat et se montre sensible à des variations d’effectifs ou de disponibilité et de qualité des
ressources peut être potentiellement considérée comme un ICE. Le principe théorique sous-jacent est
le processus de densité-dépendance: à un certain niveau de densité, les ressources disponibles pour
un individu donné diminuent, ce qui est susceptible d’entraîner une série de modifications biologiques
(baisse de la masse corporelle, augmentation de l’impact sur le milieu,…).
Il reste aujourd’hui à coordonner et spécifier le cadre d’utilisation de ces outils. Tout d’abord, seule la
multiplicité des paramètres utilisés permet de caractériser et d’interpréter les changements d’état du
système “individu-population-environnement”. Là où les dénombrements ne s’intéressent qu’à l’effectif
des populations, les ICE s’appuient sur trois composantes distinctes: l’abondance de la population, la
performance individuelle et l’impact des animaux sur l’habitat. En effet, travailler sur l’abondance seule
ou n’utiliser qu’un type d’ICE n’a pas de sens ! On peut l’illustrer avec un exemple simple. Sur la
figure 3a, on ne dispose que d’un suivi d’abondance qui ne fournit donc aucune information sur la
relation entre la population et son habitat. Avec les 3 indicateurs, il est par contre possible de savoir si
notre population est plutôt stable (aucune variation sensible des indicateurs au cours du temps,
figure 3b) ou si la situation est plus problématique (figure 3c). On est typiquement pour la figure 3c
dans une situation où le niveau de ressource n’est plus en adéquation avec le nombre d’individus
présents. On pourrait alors préconiser, par exemple, la stabilité du plan de chasse pour le scénario de
la figure 3b et l’augmentation du plan et/ou des ressources disponibles pour le scénario de la
figure 3c. Ainsi, un même constat de stabilité de l’effectif peut donc conduire à deux mesures de
gestion complètement opposées qu’il n’aurait pas été possible de distinguer sans avoir recours à une
batterie d’indicateurs ! La mesure de ces paramètres n’a donc vraiment de sens qu’au travers de
leur complémentarité, inscrite dans un suivi temporel de longue durée.
Enfin, il ne faut pas perdre d’esprit que l’utilisation de ces ICE repose sur un certain nombre
d’hypothèses. Dans notre exemple (figure 3), l’indicateur abondance doit réellement réagir aux
variations d’effectifs de la population et les indicateurs performance et habitat aux variations de
densité (effectif par rapport aux ressources) pour pouvoir être qualifiés d’ICE. Valider scientifiquement
une mesure ou une méthode de suivi comme ICE constitue l’objectif actuel d’une partie des
recherches menées par l’ONCFS et le CNRS et c’est l’état de nos connaissances dans le cadre du
suivi des populations de Cerfs que nous allons maintenant aborder (voir aussi la communication de
Christophe Bonenfant lors de ce colloque).
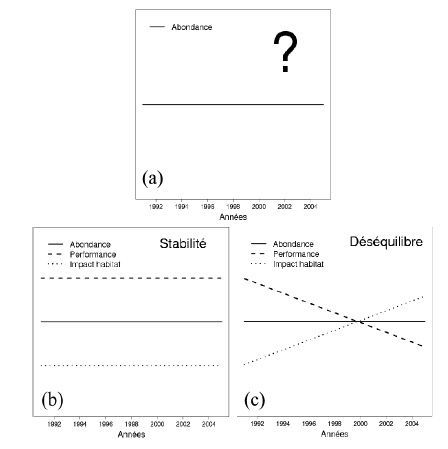
Figure 3 Exemple théorique d’une gestion par ICE. Sur la figure (a), le gestionnaire dispose uniquement d’un
suivi d’abondance qui ne lui permet pas de statuer sur l’état de son système population–
environnement. En ajoutant le suivi de deux autres composantes, l’habitat et la performance
individuelle, le gestionnaire sera alors capable de trancher entre différents scénarios
“démographiques” et pourra ainsi prendre les décisions de gestion adaptées aux objectifs qu’il s’était
fixé.
Applications pratiques: le Cerf de la Petite Pierre
Une réserve, des captures, de la chasse
Les résultats présentés ici proviennent des suivis réalisés sur la population de cerfs de la réserve
nationale de chasse et de faune sauvage de la Petite Pierre (67). Ce site d’étude de l’ONCFS, co-géré
avec l’ONF, offre un contexte de travail particulièrement adapté à la mise au point d’ICE. Cette
population à l’origine “source” d’animaux pour la ré-introduction de cerfs en France a subit une
diminution marquée de ses effectifs lorsque le maintien d’importante densité s’est avéré incompatible
avec les objectifs sylvicoles. La population est ainsi passée par une première période à “forte” densité
puis par une période de plus faible densité (figure 4) qui est maintenue depuis une vingtaine d’années.
Ces conditions fournissent les moyens d’évaluer la sensibilité de différents indicateurs aux variations
de densité.

Figure 4 Variation de l’abondance de la population cerfs de la Petite Pierre sur la période 1977–1999. La
population est d’abord passée par une phase de haute densité (population source pour les
réintroductions du cerf en France), puis par une phase de basse densité suite à l’augmentation du plan
de chasse afin de satisfaire les objectifs sylvicoles.
En outre, cette population fait l’objet de nombreux suivis depuis les années 70: analyses du tableau
de chasse et suivi de l’abondance à travers l’utilisation de méthodes comme les comptages aux
phares. Un suivi longitudinal et continu par capture-marquage-recapture d’animaux a aussi été mis en
place. Toutes ces informations permettent à la fois de disposer de nombreuses mesures pouvant
jouer le rôle d’ICE et d’avoir une connaissance détaillée du fonctionnement démographique pour
tester la pertinence de ces potentiels candidats comme ICE.
ICE abondance: les comptages phares à la Petite Pierre
Pourquoi s’intéresser aux comptages aux phares ? Tout d’abord parce que c’est une méthode très
largement utilisée dans le monde de la gestion. En effet, c’est un suivi qui “motive” puisqu’il prend
place de nuit et permet d’observer une grande diversité faunistique. Cependant à l’heure d’aujourd’hui
très peu d’informations sont disponibles sur la pertinence d’un tel suivi.
Dans le cadre des suivis réalisés à la Petite Pierre, nous disposons de 768 comptages aux phares
réalisés simultanément sur 3 circuits. Les circuits sont parcourus depuis 1978 en réserve de 3 à 11
fois par an sur la période de décembre–avril. Ces suivis ont permis de réaliser 15459 observations
dont 1082 observations d’animaux marqués. Les observations des animaux marqués ont été utilisées
chaque année pour obtenir une estimation de l’effectif de cerfs présents grâce à la méthode
d’Arnason-Schwartz-Gerrard (Arnason et al. 1991). Cette estimation de l’effectif qui n’est possible que
par la présence d’animaux marqués (donc identifiables) dans la population a été confrontée à
différents indices d’abondance, dont le nombre moyen d’animaux recensés par sortie une année
donnée.
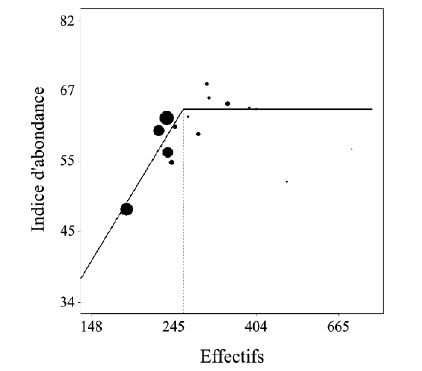
Figure 5 Relation entre le nombre moyen d’animaux observés lors du suivi aux phares de la population de
cerfs de la Petite Pierre et son effectif estimé par capture-marquage-recapture. La droite représente la
relation linéaire jusqu’à environ 260 individus puis la saturation entre ces deux mesures. La taille des
points est proportionnelle à la précision de l’estimation d’effectif. Plus le point est important, plus la
précision est bonne.
Les résultats obtenus (figure 5) sont très prometteurs puisque jusqu’à un certain seuil (~260 cerfs)
l’indice réagit de manière linéaire aux variations d’effectifs et constitue à ce titre un bon candidat
comme ICE d’abondance. Au delà de ce seuil l’indice n’augmente plus lorsque l’effectif augmente,
phénomène de saturation classique des indices basés sur l’observation d’animaux. Autres résultats
intéressants dans un cadre de gestion: il est préférable d’utiliser comme indice d’abondance le
nombre moyen d’animaux observés plutôt que le nombre maximal.
Quelques conseils lors de l’utilisation des suivis d’abondance
Comme nous l’avons étudié précédemment, nous ferons référence ici aux méthodes indiciaires
d’abondance (voir Cugnasse & Garel 2003 pour plus de détails), c’est à dire aux méthodes de suivi
basées sur des indices d’abondance et dont l’objectif n’est pas d’obtenir l’effectif total de la population
(approches qui doivent être définitivement oubliées) mais simplement un indice qui est corrélé à cet
effectif. Notre indice représente un certaine proportion de l’effectif observable sur la surface
échantillonnée. Cette proportion, c’est notre probabilité de détecter les animaux présents. Si celle-ci
est de 1, alors notre indice est égal au nombre d’animaux présents sur la zone échantillonnée. On sait
cependant qu’il est difficile de détecter tous les animaux présents. Notre probabilité de détection est
donc souvent inférieur à 1. Mais cela ne posera aucun problème tant que cette probabilité de
détection reste constante au cours du temps. Illustrons ce principe en prenant l’exemple d’un effectif
stable, égal à 100, sur la zone échantillonnée. Si une année donnée notre probabilité de détection est
égale à 0.6, alors nous verrons 60 animaux durant le suivi. Si l’année suivante cette probabilité est
égale à 0.4, nous recenserons 40 individus. Les variations de probabilité de détection nous donnerons
alors la fausse impression que l’abondance de la population a diminué au cours du temps, puisque
notre indice d’abondance diminue (60 puis 40), alors que l’effectif est resté stable (100) ! Il faut donc
que la probabilité de détection soit constante au cours des suivis de sorte que si l’indice diminue c’est
bien parce qu’il y a moins d’individus, et non pas parce que nous les détectons moins bien (et ce pour
n’importe quelle raison: par exemple une effet observateur). Cela peut paraître trivial, mais c’est un
point largement ignoré !
Pour le gestionnaire, la règle qui découle de tout ça est relativement simple: contrôler pour tout
facteur susceptible de faire varier la probabilité de détection. Il est possible de contrôler directement
les variations de probabilité de détection avec la mise en place d’un protocole adapté (choix des
observateurs, des périodes d’observations,…); mais aussi indirectement au cours des analyses. A
titre d’exemple, l’indice d’abondance obtenu sur la réserve de la Petite Pierre diminue de 17% entre
les sorties où la visibilité est bonne et les sorties où elle est moyenne. Il y a deux solutions ici:
supprimer les sorties réalisées dans de moins bonnes conditions ou (solution choisie dans notre cas)
corriger dans l’analyse l’indice par ce facteur “visibilité”.
ICE performance: le suivi du poids et du taux de femelles gestantes
L’information provient ici de l’analyse du tableau de chasse (pour plus de détails voir Bonenfant et al.
2002). Les données récoltées par les chasseurs nous ont permis de disposer du poids éviscéré des
animaux prélevés et du statut de gestation des femelles (analyse de leur tractus génital). On s’attend
évidemment à ce que le poids et la proportion de femelles gestantes augmentent en période de faible
densité (plus de ressources par individu) qu’en période de forte densité (Gaillard et al. 2000); et c’est
bien ce que l’on observe sur les figures 6 et 7. Les variations avec la densité de ces deux paramètres
valident clairement leur utilisation comme ICE performance. Il est par ailleurs cohérent que ces deux
paramètres montrent le même signal dans la mesure où l’on sait que la reproduction chez les ongulés
est étroitement lié à la masse corporelle (Sadleir 1987). On observe enfin que la “réaction” est plus
marqué chez les jeunes que chez les adultes. Chez une espèce comme le cerf ce résultat était
attendu (Clutton-Brock et al. 1982) et préconise de suivre en priorité le poids des jeunes et la
proportion de bichettes gestantes afin d’être capable de détecter plus finement des variations de
densité.

Figure 6 Poids éviscéré (kg) des femelles prélevées à la chasse durant les périodes de haute et basse
densités, respectivement. Les résultats obtenus chez les mâles sont qualitativement similaires.

Figure 7 Proportion de femelles gestantes durant les périodes de haute et basse densités, respectivement.
Quelques conseils pour l’analyse du tableau de chasse
Comme pour les suivis d’abondance, utiliser des ICE performance ne peut se faire sans un minimum
de précaution. Parmi les points qui nous paraissent important, il y a la nécessité de corriger les
mesures de poids par la date de récolte. En effet, que ce soit les jeunes individus qui ont une
croissance importante durant leur première année de vie ou les adultes qui peuvent perdre du poids
pendant la période d’accouplement, il faut prendre en compte ces variations de poids pour rendre
comparable les mesures réalisées à différentes périodes de l’année. Sur la figure 8, le faon 2 (poids 2)
semble plus lourd que le faon 1 (poids 1). Tout deux ont été prélevés en période de chasse alors qu’ils
sont encore en croissance comme indiqué par la droite continue. On corrige le poids de ces animaux
et on le ramène à une date donnée, par exemple le premier octobre. L’idée ici est notamment de
prendre en compte la croissance qu’aurait eu l’animal 1 si on lui avait laissé le temps de grandir
jusqu’au premier octobre. Avec cette correction, on constate en réalité que le faon 1 est plus gros que
le faon 2 à une date donnée.

Exemple théorique du bien fondé de corriger le poids des animaux par la date de mesure. Sans
correction, l’animal 1 semble plus léger que l’animal 2. Après correction, il s’avère que c’est la
situation inverse qui est vraie.
En ce qui concerne le calcul de la proportion de femelles gestantes, il faut bien garder à l’esprit que
chez le cerf les femelles peuvent avoir plusieurs oestrus et ne sont pas forcément fécondées dès le
premier (Sadleir 1987). Pour être sûr qu’une femelle n’est pas gestante, il est donc préférable de la
prélever le plus tard dans la saison de chasse, après la date supposée de fin de période
d’accouplement. C’est uniquement sur cet échantillon de femelles qu’il faudra calculer la proportion de
celles qui sont gestantes.
Perspectives et conclusions
Il existe encore aujourd’hui un grand nombre de méthodes de suivi des populations de cerfs qui n’ont
pas fait l’objet d’une évaluation scientifique rigoureuse et qui ne peuvent donc être reconnu comme
ICE. Les ICE renseignant sur l’habitat font à ce titre encore défaut. Il faut donc poursuivre l’évaluation
des méthodes existantes en s’appuyant notamment sur des sites de référence comme celui de la
Petite Pierre. La manipulation des prélèvements fournit des conditions favorables d’évaluation des
différents indicateurs comme nous l’avons vu pour les comptages aux phares. Dans la mesure du
possible, il serait particulièrement bénéfique de développer ce genre d’approche.
En définitive, les indicateurs population/environnement constituent les méthodes d’avenir pour le suivi
des populations d’ongulés en France. Simple d’emploi, ils prennent en compte l’animal dans son
environnement et permettent de rechercher un équilibre entre ces deux composantes en fonction des
objectifs définis préalablement par le gestionnaire. Plutôt que de caractériser une situation ponctuelle,
ils informent sur l’histoire et l’évolution du “système” et permettent ainsi de faire une gestion
raisonnée, a posteriori, des populations.
Bibliographie
Andersen, J. 1953. Analysis of a danish roe-deer population (Capreolus capreolus (L)) based upon the
extermination of the total stock. Danish Review of Game Biology 2 : 127–155.
Arnason, A.N., Schwarz, C.J. & Gerrard, J.M. 1991. Estimating closed population size and number
of marked animals from sighting data. Journal of Wildlife Management 55 : 718–730.
Bonenfant, C., Gaillard, J.M., Klein, F. & Loison, A. 2002. Sex- and age-dependent effects of
population density on life-history traits of red deer Cervus elaphus in a temperate forest. Ecography,
25 : 446–458.
Boscardin, Y. 1999. Les méthodes de dénombrement des populations de chevreuils. Bulletin Mensuel
de l’Office National de la Chasse 244 : 17–21.
Bourlière, F. 1969. Les techniques d’échantillonnage utilisables pour l’étude des populations de
grands mammifères sauvages. Revue d’Ecologie-La Terre et la Vie 2 : 238–244.
Chardonnet, P., des Clers, B., Fischer, J., Gerhold, R., Jori, F. & Lamarque, F. 2002. The value of
wildlife. Revue scientifique et technique de l’office international des epizooties 21 : 15–51.
Clutton-Brock, T.H., Guinness, F.E. & Albon, S.D. 1982. Red deer. Behaviour and ecology of two
sexes. Edinburgh University Press, Edinburgh, UK.
Cugnasse, J.M. & Garel, M. 2003. Suivi de l’abondance des populations d’Ongulés sauvages en
montagne : l’exemple du Mouflon méditerranéen. Faune Sauvage 260 : 42–49.
Gaillard, J.M. 1988. Contribution a la dynamique des populations de grands mammifères : l’exemple
du chevreuil (Capreolus capreolus). Ph.D. thesis, Université Claude Bernard Lyon 1, Lyon, France.
Gaillard, J.M., Festa-Bianchet, M., Yoccoz, N.G., Loison, A. & Toïgo C. 2000. Temporal variation in
fitness components and population dynamics of large herbivores. Annual Review of Ecology and
Systematics 31 : 367–393.
Lecocq, Y. & Meine, K. 1998. Hunter demography in Europe - An analysis. Gibier Faune Sauvage 15 :
1049–1061.
Morellet, N., Gaillard, J.M., Hewison, A.J.M., Ballon, P., Boscardin, Y., Duncan, P., Klein, F. &
Maillard, D. 2007. Indicators of ecological change : new tools for managing populations of large
herbivores. Journal of Applied Ecology 44 : 634–643.



/image%2F1498946%2F20150921%2Fob_75a45c_la-pilotie-re-ouverture-2015-009.jpg)
/http%3A%2F%2Fs2.dmcdn.net%2FOQGtv%2Fx240-wN2.jpg)
/idata%2F3490124%2F20131013.ChasseADM%2FIMGP0768.JPG)
/idata%2F3490124%2F2013.09.28.Chasse-Pilotiere%2FIMGP0646.JPG)